Comparación del tamaño y la capacidad de germinación de semillas dispersadas por Alouatta seniculus en paisajes con diferentes grados de fragmentación, durante la época de lluvias en San Martín, Meta.Daniela Gaitán Gómez Pontificia Universidad Javeriana 2017
La especie con mayor frecuencia y abundancia en cada paisaje fue Bellucia grossularoides. Se presentaron diferencias en los tamaños de las semillas encontradas en cada paisaje. No se presentaron diferencias en términos de capacidad de germinación entre los paisajes, pero sí hay diferencias entre los tiempos para cada especie de semilla encontrada. Adicionalmente se evaluaron la similitud en términos de sobreposición y amplitud de dieta además del índice de importancia de cada especie de planta para cada paisaje. ◼
Contacto: gaitan.daniela@javeriana.edu.co. Conoce cómo publicar el resumen de tu tesis en la Red-Primatológica ingresando aquí. Por Jeisson Castro Salamanca Consejo Directivo APC En la última década, el tití gris (Saguinus leucopus) se ha convertido en una de las cinco especies más estudiadas de Colombia, y en la segunda especie con mayor número de publicaciones enfocadas en temas de conservación, después del tití cabeciblanco, Saguinus oedipus. El evidente progreso en el estudio de esta especie, y las numerosas publicaciones sobre aspectos relacionados con su conservación, son el resultado de diferentes iniciativas y proyectos que han contribuido de forma significativa y efectiva al conocimiento y la protección de sus poblaciones.  Saguinus leucopus. Foto: Camilo Botero, La Bici Errante. Uno de los principales logros para la conservación del tití gris ha sido el desarrollo y publicación en el 2008 del “Programa Nacional para la Conservación de la especie endémica de Colombia, Tití gris (Saguinus leucopus)”, que contó la participación de varios actores, incluyendo a CORPOCALDAS y el Ministerio de Ambiente y Desarrollo Sostenible. En este programa se elaboró un modelo de distribución potencial de la especie mediante herramientas SIG, que fue evaluado con respecto a la pérdida de cobertura de bosque en el área de distribución. Para esto se usaron los mapas de ecosistemas de Colombia y el mapa de bosques del IDEAM, y 69 localidades obtenidas de diferentes fuentes.
Gracias a otras iniciativas y proyectos como el “Programa Internacional de Conservación de Saguinus leucopus”, y el “Plan de conservación y manejo del Tití gris (Saguinus leucopus) del Sistema Regional de Áreas Protegidas del Eje Cafetero Colombiano”, y al trabajo de entidades como Wildlife Conservation Society Colombia (WCS-Colombia), la Asociación Colombiana de Parques Zoológicos y Acuarios (ACOPAZOA), Conservación Tití Gris y la Universidad de Antioquia, entre otras; el tití gris fue la única especie colombiana que, tras la evaluación del estado de conservación de los primates neotropicales realizada por el Grupo de Especialistas en Primates de la UICN/CSE en enero de 2015, bajó de categoría de amenaza, pasando de ‘En Peligro’ (EN) a ‘Vulnerable’ (VU). No obstante, pese a los avances logrados, es necesario continuar fortaleciendo los esfuerzos por la conservación de este primate endémico de Colombia. En particular, es fundamental mejorar la cobertura de su distribución por parte del Sistema Nacional de Áreas Protegidas. La creación del PNN Serranía de San Lucas favorecería la conservación no solo del tití gris, sino también al mono araña del magdalena (Ateles hybridus), una de las cuatro especies de primates colombianos clasificadas como ‘En peligro crítico’ (CR). ◼ REFERENCIAS
Por Jeisson Castro Salamanca Consejo Directivo APC Los huapos rojos o uakaris calvos (Cacajao calvus) son primates neotropicales que se distribuyen a lo largo de las zonas pantanosas de la Amazonía peruana y brasileña. Su apariencia física es bastante particular, ya que no presenta pelaje alrededor de su rostro pero sí una coloración roja bastante intensa. En un estudio publicado en el 2016, investigadores de la Universidad Federal de Rio Grande do Sul, del Instituto de Conservación del Zoológico de San Diego, el Instituto de investigación Leibniz y la Universidad de Cambridge, encontraron otra particularidad de los huapos rojos y es que éstos presentan una visión del color altamente polimórfica. Esto quiere decir que la visión del color varía ampliamente entre individuos, donde algunos presentan visión dicromática, común en la mayoría de especies de primates neotropicales, y otros visión tricromática, ambas asociadas a seis alelos funcionales.  Cacajao calvus. Foto tomada de: ARKIVE - Thomas Marent Existen diferentes hipótesis que pueden explicar esta alta variabilidad. Entre ellas, se cree que la visión tricromática está más asociada a procesos de selección sexual, donde la coloración roja del rostro resulta ser un buen indicador de la salud de los machos. Por otra parte, la visión dicromática está más asociada a procesos de selección de frutos, ya que estos primates tienen una dieta rica en frutos verdes, especialmente crípticos, y este tipo de visión puede ser de gran ayuda para la identificación de éstos. ◼
Fuente: Corso et al. 2016. Highly polymorphic colour vision in a New World monkey with red facial skin, the bald uakari (Cacajao calvus). Proceedings of the Royal Society B: Biological Sciences 283 (1828). Ver publicación. Notas relacionadas La visión en primates: No todos vemos el mundo del mismo color.  Por Gianna Cristhina Flórez Ex-voluntaria APC Los patrones fenológicos de las plantas determinan la disponibilidad de alimento en el tiempo para numerosas especies de primates. En consecuencia, el estudio de estos patrones es una importante herramienta para conocer diversos aspectos de la ecología de estas especies. En particular, los estudios fenológicos permiten explorar en detalle la relación entre diferentes aspectos de la dieta y el comportamiento de las poblaciones de primates, y los patrones de productividad de los bosques que habitan. Por esto, este tipo de estudios no solo contribuyen significativamente a la comprensión de los mecanismos reproductivos de las plantas [1], sino que también contribuyen al conocimiento de las interacciones planta-animal, la dieta y las preferencias alimenticias de frugívoros. El estudio de los ciclos de crecimiento y reproducción de las plantas, y de las variables que los afectan es particularmente importante para entender la ecología de los primates frugívoros [2]. Al determinar la distribución espacio-temporal de su principal alimento, los patrones fenológicos influyen sobre aspectos tan importantes de estos primates como el uso de hábitat, el tamaño de los rangos de hogar, la longitud recorridos diarios, preferencia alimenticia, y las dinámicas de competencia intra- e inter-específica [3, 4, 5, 6]. Por esto, la integración de información fenológica a los estudios sobre éstos y otros aspectos de la ecología de los primates frugívoros es crucial para conocer, más allá de un plano descriptivo, sobre las adaptaciones y respuestas de estas especies a las dinámicas de su ecosistema.
Por ejemplo, la productividad anual de frutos y la estacionalidad en la producción se han establecido como elementos importantes determinando la abundancia de los ensamblajes de primates [9, 10], y justamente las poblaciones poco densas son las que tienen mayor chance de que surjan extinciones. Fenología de PlantasLa fenología de plantas involucra la observación, registro e interpretación de eventos tales como la producción de hojas, flores y frutos; y el estudio de los factores bióticos y abióticos que los ocasionan o los afectan. Hay cuatro formas principales para estudiar las fases fenológicas. La primera es utilizar trampas de frutos (e.g. rectángulos de tela porosa de área conocida) [11, 12], monitorear las plantas a lo largo de transectos o líneas de muestreo [13], revisar todos los individuos en parcelas de vegetación de área conocida [14] y marcar varios (ca. 6-10) individuos de las especies de interés [15].  Siphocampylus scandens (Campanulaceae) en PNN Cueva de Los Guácharos. Foto: LEBTYP. Para efectos de cuantificar la producción total de una comunidad, estudios recientes han sugerido que la mejor metodología es la de trampas de frutos, sin embargo, es importante recalcar que se requiere al menos 100 trampas por tipo de bosque, e idealmente 200 o 300 [16]. Los muestreos por transectos fenológicos son adecuados para mostrar los momentos en que diferentes especies producen frutos o flores, y han sido utilizados en análisis de preferencias alimenticias [17]. El esfuerzo de muestreo requerido para un buen muestreo por transectos depende de la diversidad de los bosques; así, se han utilizado longitudes totales de 10 km en bosques muy diversos [8] o de 3 km en ecosistemas menos diversos [15]. Los patrones fenológicos incluyen el momento de ocurrencia, la frecuencia, duración y grado de sincronía de cada fase [18, 19]. Por ejemplo, en bosques neotropicales la producción de flores tiende a ser mayor en épocas secas, lo que puede estar relacionado con mayor actividad de insectos polinizadores [1]; mientras que la producción de nuevas hojas suele ser más común al final éstas épocas, cuando hay menos insectos herbívoros [20]. El momento de producción de frutos depende del tipo de dispersión de semillas. Frutos dispersados por viento se producen en épocas secas, que suelen presentar fuertes viento; mientras que aquellos dispersados por animales (como primates), se producen mayormente al comienzo de la época de lluvias. Esto con el fin de que las semillas puedan germinar rápidamente, y así las plántulas tengan un suministro de agua durante varios meses (hasta la siguiente época seca).
Se ha sugerido que la sincronización se logra debido a cambios climáticos en temperatura, que están asociados a eventos de El Niño. Este mecanismo no es tan común en el Neotrópico, posiblemente porque las plantas van a dispersar más semillas si mantienen altas las poblaciones de dispersores, al producir frutos y semillas con mayor regularidad. De hecho, en lugares con suelos fértiles, las mayoría de las especies producen frutos una o dos veces al año [1], mientras que en suelos poco fértiles son supra-anuales y anuales [22], ya que la fertilidad del suelo y la irradiación limitan la producción de frutos [12]. Por lo tanto, se puede concluir que los patrones de producción de flores y frutos son el resultado de la interacción entre variables genéticas, fisiológicas y climáticas. Adicionalmente, estos patrones pueden ser afectados por la disponibilidad de recursos, por las interacciones de competencia, y por las interacciones con otros organismos tales como polinizadores, depredadores o dispersores de semillas.
REFERENCIAS[1] Stevenson et al. 2008. Flowering patterns in a seasonal tropical lowland forest in western Amazonia. Biotropica 40(5): 559-567.
[2] van Schaik et al. 1993. The phenology of tropical forests: Adaptive significance and consequences for primary consumers. Annual Review of Ecology and Systematics 24: 353-377. [3] Peres 1994. Primate responses to phenological changes in an Amazonian terra firme forest. Biotropica 26(1): 98-112. PDF [4] Janson & Goldsmith 1995. Predicting group size in primates: Foraging costs and predation risks. Behavioral Ecology 6: 326-336. [5] Stevenson et al. 2000. The relationship between temporal variation in fruit abundance and ecological overlap among four neotropical primates in Colombia. Biotropica 32(3): 533-544. [6] Stevenson & Castellanos 2001. Feeding rates and daily path range of the Colombian woolly monkeys as evidence for between-and within-group competition. Folia primatologica 71(6): 399-408. [7] Chapman et al. 2005. A 12-year phenological record of fruiting: Implications for frugivore populations and indicators of climate change. En: Tropical Fruits and Frugivores. The search for strong interactors (JL DEW, JP BOUBLI eds.) Springer. PDF [8] Vargas & Stevenson 2009. Patrones fenológicos en la Estación Biológica Mosiro Itajura-Caparú: producción de frutos estimada a partir de transectos fenológicos y trampas de frutos. En: Estación Biológica Mosiro Itajura-Caparú, Biodiversidad En El Territorio Del Yaigojé-Apaporis. Conservacion Internacional Colombia, pp: 99-114. [9] Stevenson 2001. The relationship between fruit production and primate abundance in Neotropical communities. Biological Journal of Linnean Society 72: 161-178. PDF [10] Hanya et al. 2011. Seasonality in fruit availability affects frugivorous primate biomass and species richness. Ecography 34: 1009-1017. PDF [11] Terborgh 1983. Five new world primates: a study of comparative ecology. Princeton University Press, Princeton. [12] Wright et al. 1999. The El Niño Southern Oscillation, variable fruit production, and famine in a tropical forest. Ecology 80: 1632–1647. [13] Stevenson 2004. Fruit Choice by Woolly Monkeys in Tinigua National Park, Colombia. International Journal Of Primatology 25: 367-381. [14] Blake et al. 1990. Quantifying abundance of fruits for birds in tropical habitats. Studies in Avian Biology 13:71-77. [15] Palma et al. 2011. Use of space, activity patterns, and foraging behavior of red howler monkeys (Alouatta seniculus) in an Andean forest fragment in Colombia. American Journal of Primatology 73(10):1062-71. [16] Stevenson & Vargas 2008. Sample size and appropriate design of fruit and seed traps in tropical forests. Journal Of Tropical Ecology 24: 95-105. [17] Stevenson & Link 2010. Fruit preferences of Ateles belzebuth in Tinigua Park, northwestern Amazonia. International Journal of Primatology 31(3): 393-407. [18] Newstrom et al. 1994. A new classification for plant phenology based on flowering patterns in lowland tropical rain forest trees at La Selva, Costa Rica. Biotropica 26: 141-159 [19] Wright & Calderón 1995. Phylogenetic patterns among tropical flowering phenologies. The Journal of Ecology 83(6): 937-948. PDF [20] Aide 1988. Herbivory as a selective agent on the timing of leaf production in a tropical understory community. Nature 336: 574-575. [21] Curran & Leighton 2000. Vertebrate responses to spatiotemporal variation in seed production of mast-fruiting Dipterocarpaceae. Ecological Monographs 70(1): 101-128. [22] Norden et al. 2007. Mast Fruiting Is a Frequent Strategy in Woody Species of Eastern South America. PLoS ONE 2(10): e1079. [23] Borchert et al. 2005. Photoperiodic induction of synchronous flowering near the Equator. Nature 433: 627-629. Por Jeisson Castro Salamanca Consejo Directivo APC El género Callicebus, conocidos como zogui-zogui o monos tocones, se destaca por poseer la mayor diversidad de especies de primates neotropicales. En los últimos años este género de primates ha sido objeto de atención por la descripción de nuevas especies como el tití del Caquetá (Callicebus caquetensis) en el año 2010, el tití de Milton (C. miltoni) por Dalponte y colaboradores en Brasil (2014) y el tití marrón del río Urubamba (C. urubambensis) por Vermeer y Tello-Alvarado en Perú (2015).
Teniendo en cuenta los resultados del análisis filogenético, así como la evidencia morfológica y biogeográfica, Byrne y colaboradores sugieren una taxonomía en la que se reconocen tres géneros como parte de la subfamilia Callicebinae, que son coherentes con los resultados arrojados por el trabajo Kobayashi (1995) que se basó en mediciones craneales. 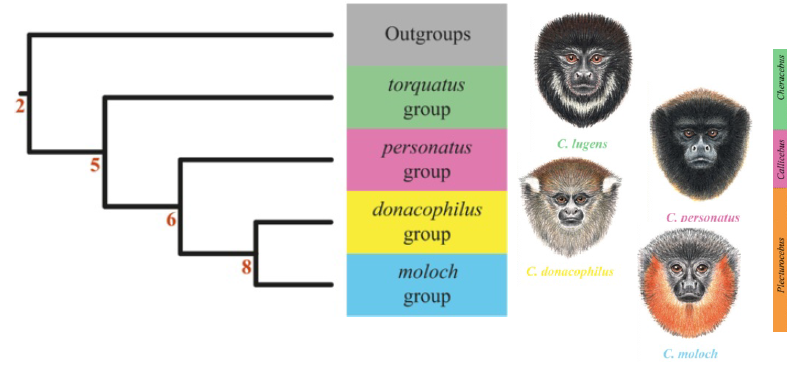 Reconstrucción filogenética de los ‘grupos de especie’ de Callicebus y el respectivo cambio de género. Tomado y adaptado de Byrne et al. 2016 ‘Phylogenetic relationships of the New World titi monkeys (Callicebus): first appraisal of taxonomy based on molecular evidence’. Al grupo torquatus, que presenta la radiación más antigua, se le asignó el género Cheracebus, y el grupo personatus quedó bajo el género Callicebus. Los complejos de especie originalmente reconocidos como cupreus y moloch no son monofiléticos, y todas las especies que anteriormente fueron reconocidas como parte del grupo cupreus fueron asignadas al grupo moloch. El grupo donacophilus y moloch muestran una relación cercana, y por ello se les asignó el género Plecturocebus. ◼ REFERENCIAS
* Esta nota fue publicada en agosto de 2015 y, por lo tanto, no incluye los últimos cambios taxonómicos propuestos para el género Callicebus.
Sin embargo, la enorme riqueza de especies de primates neotropicales no está homogéneamente distribuida entre los 19 géneros existentes. Con 34 especies, el género Callicebus es el más diverso del Neotrópico (19% de las especies de primates neotropicales), y del mundo; seguido solo por los géneros Lepilemur (Madagascar) y Macaca (Asia y África), con aproximadamente 26 y 22 especies respectivamente.
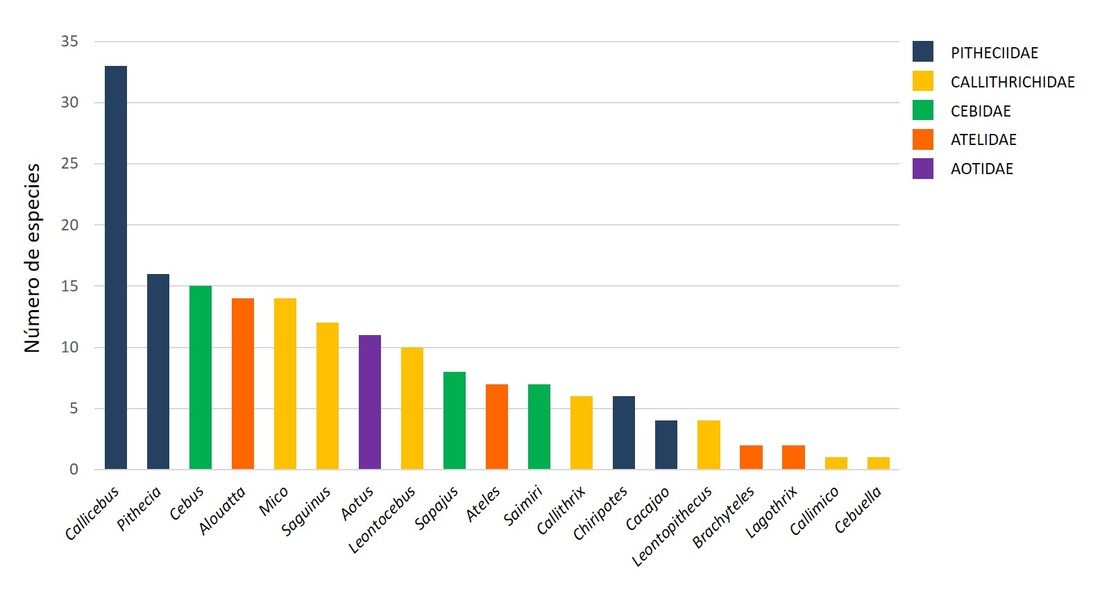
Número de especies pertenecientes a cada una de las 5 familias y 19 géneros de primates neotropicales.
En el Neotrópico, el segundo género más diverso es Pithecia (16 especies), seguido de Cebus (15 especies), Mico y Alouatta (14 especies cada uno). Por otro lado, los géneros Cebuella y Callimico (1 especie cada uno), y Lagothrix y Brachyteles (2 especies cada uno), destacan por ser los menos diversos.
Tabla. Lista de las 174 especies de primates descritas para la región neotropical.
Ayuda a mantener y proteger la gran riqueza de primates presente en nuestra región aprendiendo sobre ellos, y apoyando el trabajo de las diferentes entidades dedicadas a su estudio y conservación.
por Alma Hernández Jaramillo
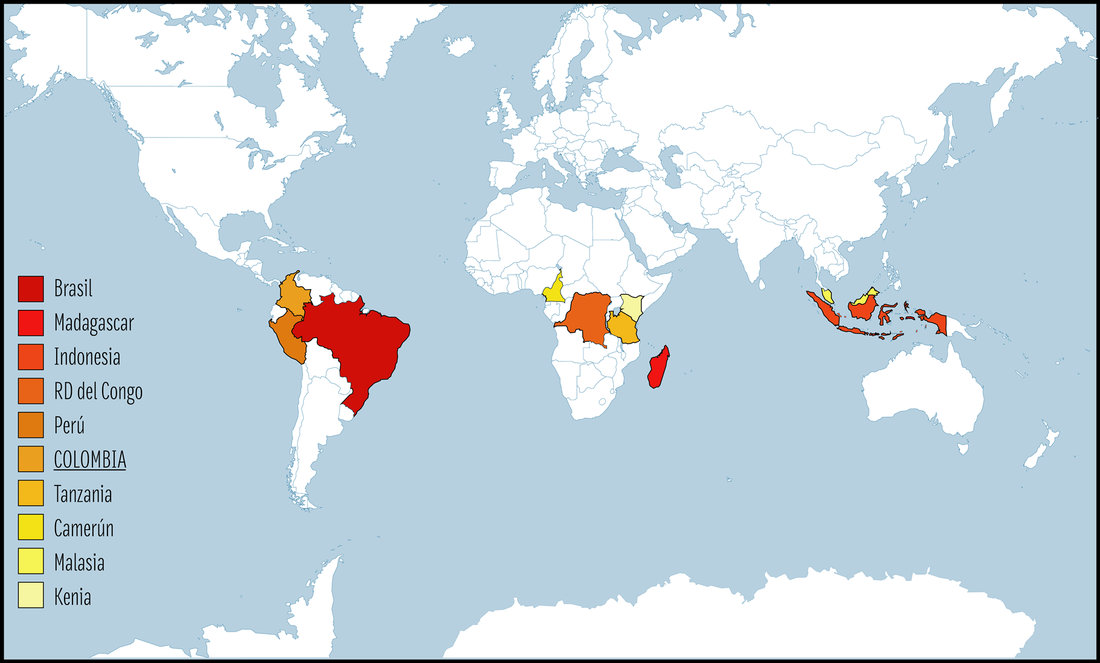
Colombia se encuentra entre los países con mayor riqueza de primates en el mundo, ocupando el sexto lugar después de Brasil, Madagascar, Indonesia, la República Democrática del Congo y Perú. De las 47 especies y subespecies presentes en Colombia, 15 no se encuentran en ninguna otra parte del mundo, en otras palabras, son endémicas de Colombia.
Las 15 especies y subespecies de primates endémicos de Colombia se agrupan en 7 géneros, pertenecientes a las 5 familias de primates presentes en el país.
La supervivencia de estos primates 100% colombianos depende en un 100% de los esfuerzos que se realicen a nivel nacional para proteger y conservar su hábitat y sus poblaciones. Por esto, la labor de organizaciones y grupos de investigación tales como la Fundación Proyecto Tití y el Laboratorio de Ecología de Bosques Tropicales y Primatología de la Universidad de Los Andes, dedicados al estudio y la conservación de primates endémicos y críticamente amenazados, es tan valiosa e importante.
Conoce más sobre los primates colombianos y las organizaciones dedicadas a su estudio y conservación a través de las publicaciones de nuestra campaña Conocer para Conservar e ingresando a los Enlaces de la RedPrim.
Los primates son un grupo de animales muy diverso. Actualmente se han descrito más de 500 especies, distribuidas en las regiones ecuatoriales de 3 continentes: África, Asia y América. Los primates se pueden dividir en 3 grandes grupos: los lémures, los simios y los monos. Los lémures se encuentran únicamente en Madagascar, los simios no-humanos habitan en África y Asia, y los monos dividen en dos grupos, los monos del Viejo Mundo (catarrinos), distribuidos en África y Asia; y los monos neotropicales o del Nuevo Mundo (platirrinos).
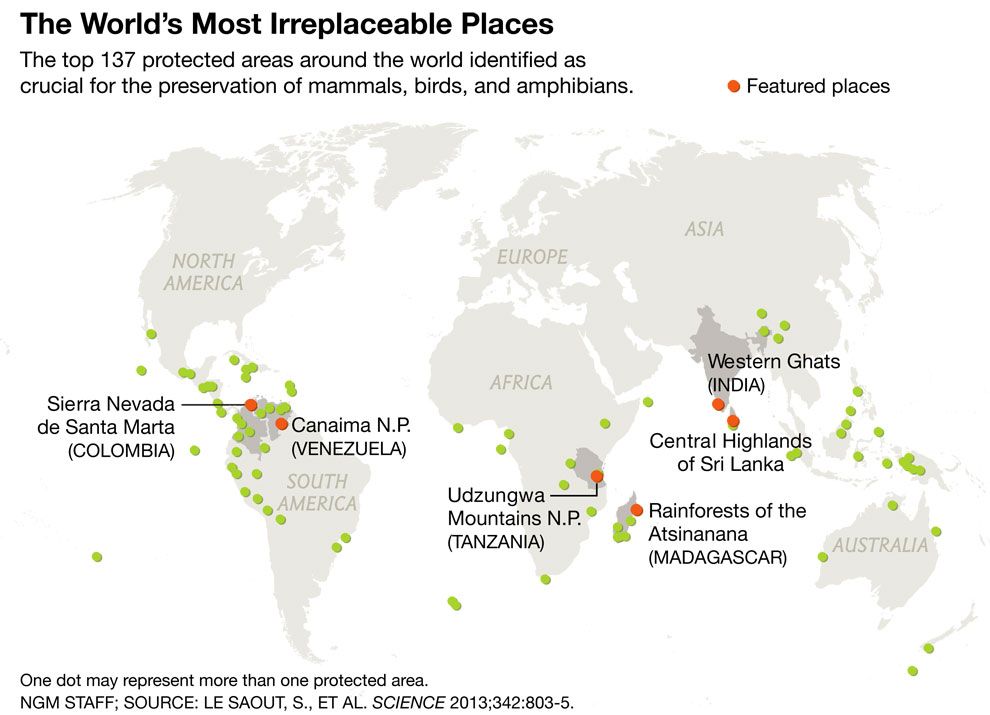
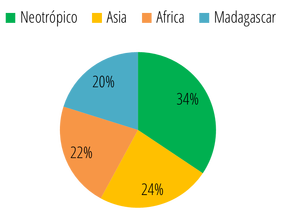
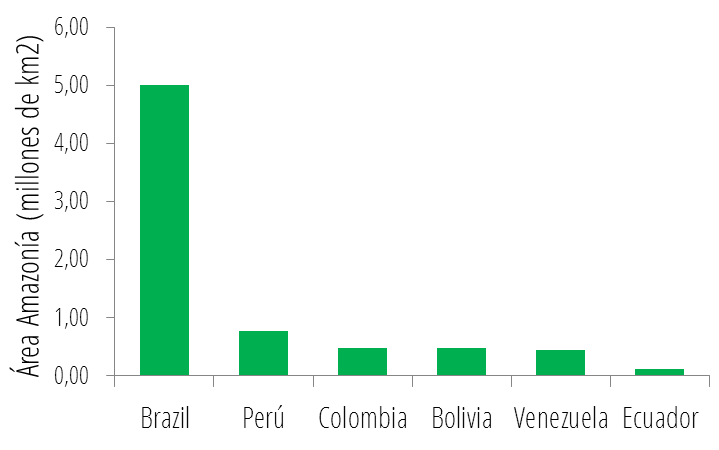
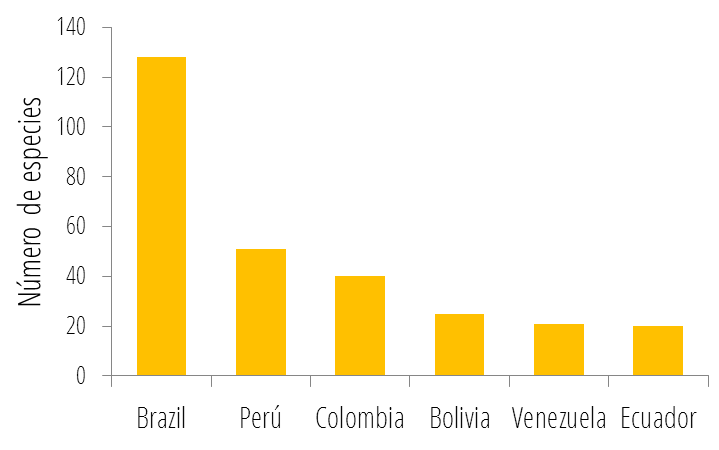
Además de Brasil, Perú y Colombia también se encuentran entre los países con mayor riqueza de primates en el mundo, con 52 y 38 especies respectivamente. La gran riqueza de primates presente en estos países, y en el Neotrópico en general, se concentra en la región de la Amazonía. Por esto existe una relación cercana entre el área de la Amazonía abarcada por los diferentes países, y el número de especies de primates distribuidas en su territorio:
De igual manera, la alta riqueza de primates en países como Indonesia en Asia, o la República Democrática del Congo en África, también está relacionada con sus grandes extensiones de bosque tropical. No obstante, los primates no habitan exclusivamente bosques tropicales, un gran número de especies habitan ecosistemas como sabanas o montañas, y no presentan hábitos arborícolas. Lamentablemente, la gran riqueza de primates del mundo se encuentra gravemente amenazada. Más de la mitad de los primates del mundo se encuentra en peligro de extinción, y un alto número de especies está clasificado como en Peligro Crítico por la Unión Internacional para la Conservación de la Naturaleza (UICN). Tan solo en el Neotrópico, la región con menor porcentaje de especies amenazadas, 70 se encuentran bajo algún grado de amenaza, 15 de ellas en Peligro Crítico: 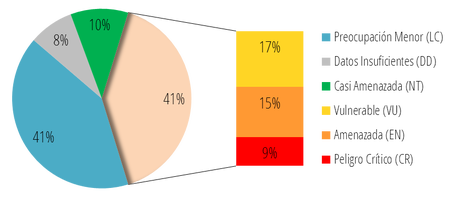 Porcentaje de especies de primates neotropicales clasificadas bajo las diferentes categorías de la UICN. De forma similar, más de la mitad de las especies de primates presentes en Colombia se encuentra en peligro de extinción, 4 de ellas clasificadas como en Peligro Crítico. Al igual que en todos los demás países hábitat, la destrucción del hábitat, la cacería y el tráfico ilegal de especies son las principales amenazas para las especies de primates en Colombia. No obstante, en los últimos años, el establecimiento de cultivos extensivos de palma de aceite y el crecimiento de la minería, se han convertido en las principales amenazas para la conservación de los primates en el país. Conoce más sobre la gran diversidad de primates del mundo y sobre cómo ayudar a su conservación, ingresando a los sitios web de las diferentes entidades dedicadas a su estudio y protección en los Enlaces de la RedPrim.
En primatología, conocer los nombres comunes de las diferentes especies es casi tan importante como conocer sus nombres científicos. El uso de los nombres comunes no sólo es la forma más frecuente de hacer referencia a las especies en conversaciones informales, sino que éstos también son usados ampliamente para identificar las especies en artículos científicos y libros.
Te invitamos a conocer algunos de los nombres comunes más usados para identificar las especies de primates colombianos, en español e inglés: |














 Canal RSS
Canal RSS

|
Asociación Primatológica Colombiana, APC
www.asoprimatologicacolombiana.org asoprimatologica@gmail.com Bogotá DC, Colombia |
Última actualización: Agosto del 2023
Proudly powered by Weebly